Major research findings involving Swiss needle cast and its causal agent:
- Fertilizer influence on SNC disease severity across a gradient of disease: The Beyond N Study
- Effects and Impacts of SNC
- SNC Treatments
- Quantification, Assessment, and Severity Prediction
- Tree Genetics, Resistance, and Improvement
All cited publications are available on our Publications page.
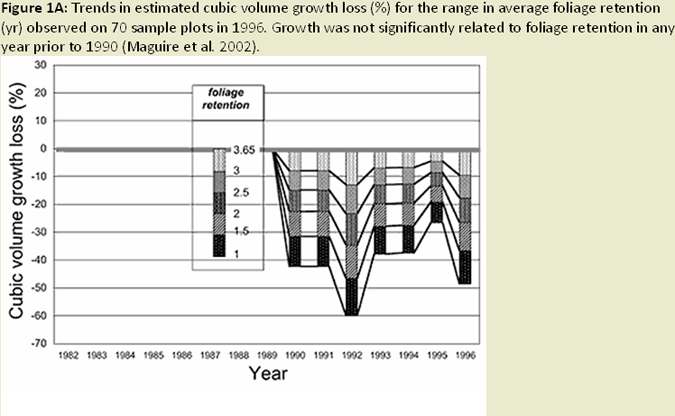
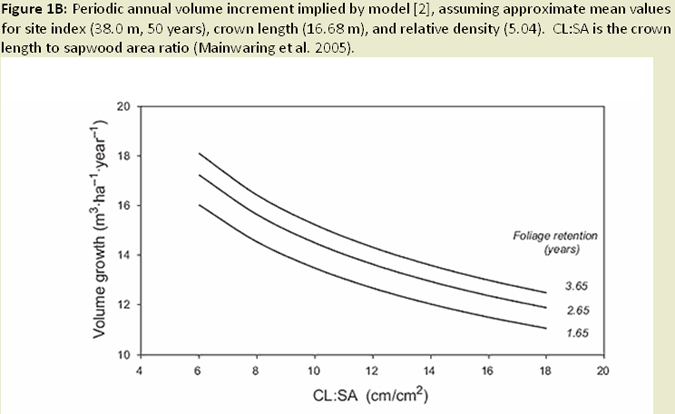
The effects of Swiss needle cast (SNC) are most severe in Douglas-fir (Pseudotsuga menziesii) plantations on coastal sites of northwestern Oregon, where Sitka spruce and western hemlock or red alder were historically dominant species (Hansen et al. 2000). General symptoms of the disease include chlorosis, decreased needle retention, and reduction in height and diameter growth. Disease impacts on height growth can reach 25%, while basal area growth losses can reach over 35% in heavily infected stands. Combined, these height and diameter losses can result in 23% (average over target population) to 52% (sites with highest infection levels) volume loss (Maguire et al. 2002; Fig. 1A; Fig. 1B). Loss of foliage was found to be the predominant mechanism reducing growth. Mainwaring et al. (2005) found similar declines in basal area growth and volume.
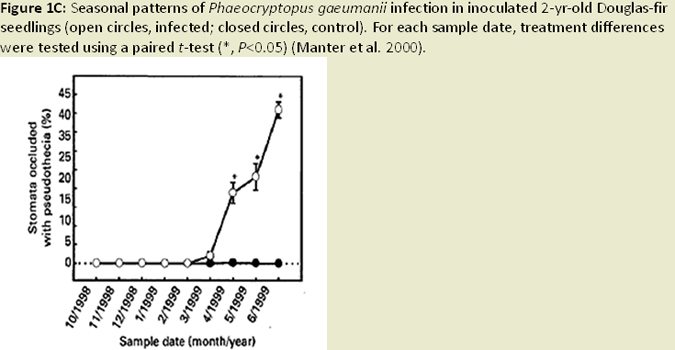
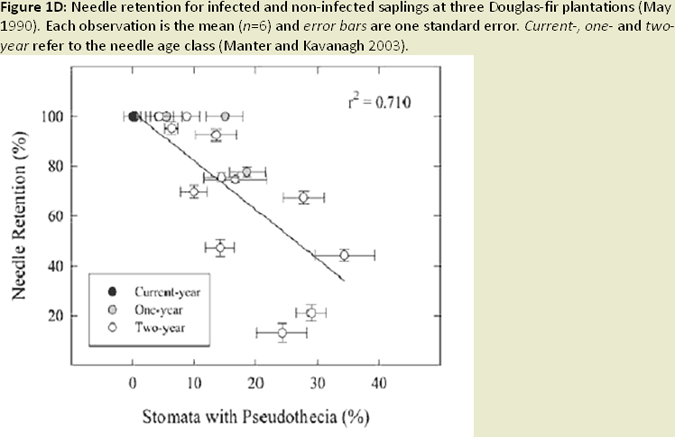
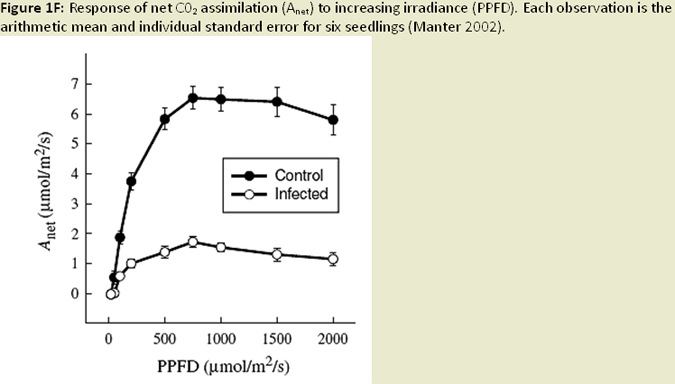
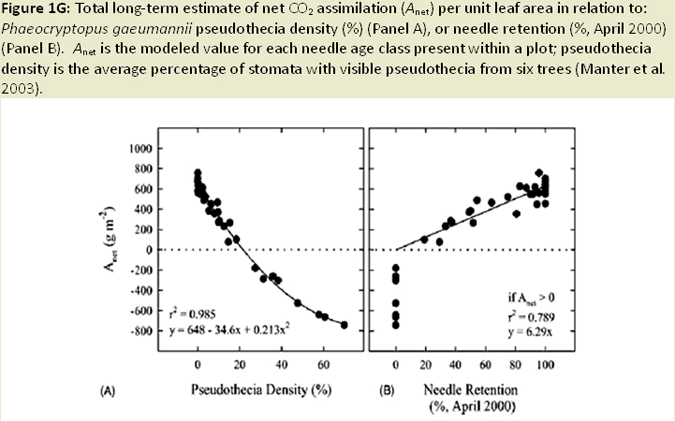
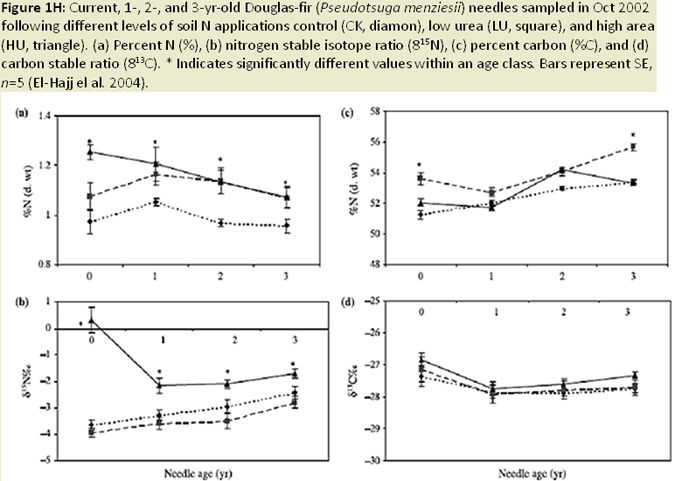
Nearly all infection occurs on newly emerging, current-season needles, and fruiting bodies (pseudothecia) emerging late in the first year west of the Oregon Coast Range (Hansen et al 2000; Fig. 1C). Needle abscission generally occurs once 50% of stomata are occluded (Hansen et al. 2000; Fig. 1D; Fig. 1E). Other studies have found that needle abscission is triggered when a certain threshold of pseudothecial density is surpassed (~25%). Stomatal occlusion impairs the needle’s ability to uptake carbon for photosynthesis, causing it to transition from a carbon source to a carbon sink (Manter et al. 2003). Models indicate that carbon assimilation (photosynthesis) in infected first year needles continues unhindered for up to 6 months, until the fungal fruiting bodies of SNC mature to a size that impairs gas exchange. Therefore, net carbon gains at the whole canopy level remain positive (Manter et al. 2003). The combination of excess absorbed light and decreased photosynthetic capacity in infected needles has been shown to cause photoxidative damage, which may also contribute to premature needle loss in sun exposed foliage (Manter 2002; Fig. 1F). Pseudothecia, emerging from needle stomata, impede gas exchange (Manter et al. 2000), reduce stomatal conductance in remaining functional foliage, (Manter and Kavanagh 2003) and decrease CO2 assimilation up to 3 fold (Manter et al. 2000; Manter 2002; Fig. 1G). The fungus also appears to gain nutrients from intercellular spaces of needles, possibly by altering membrane permeability (Hansen et al. 2000). However, the mechanism by which the fungus obtains nutrients from its host is difficult to study and is poorly understood. Studies of soil nutrient additions have demonstrated that P. gaeumannii responds to the nutrient status in host trees and that increased N-availability in Douglas-fir needles, particularly when applied as a needle surface spray, is related to the severity of SNC (El-Hajj et al. 2004; Fig. 1H).
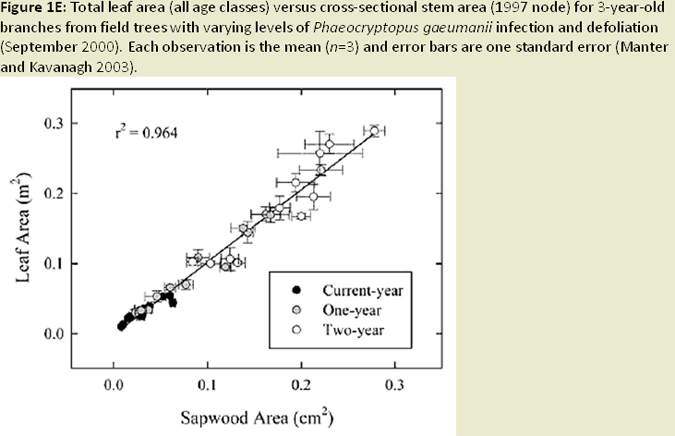
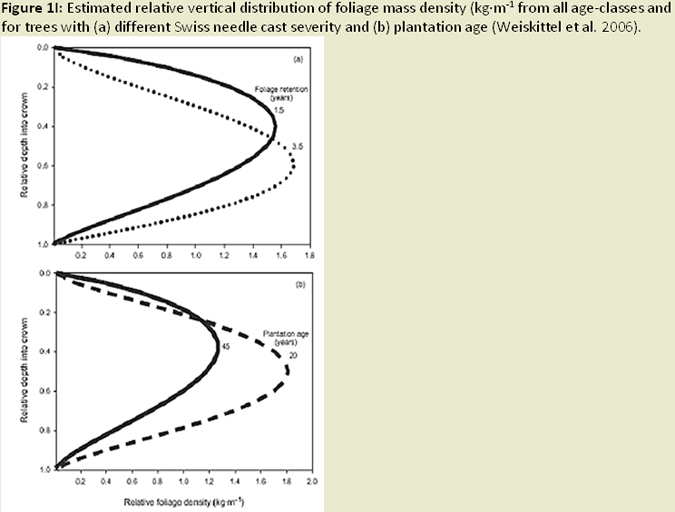
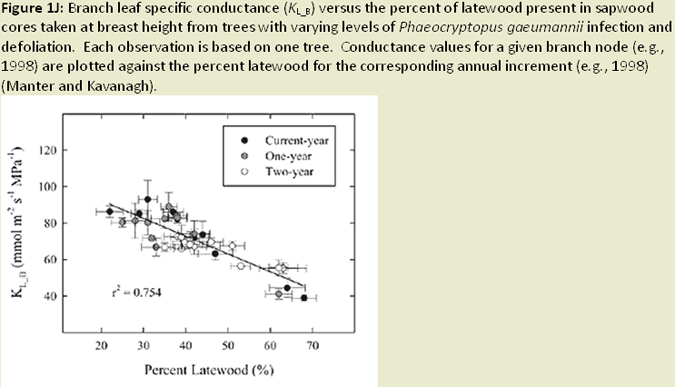
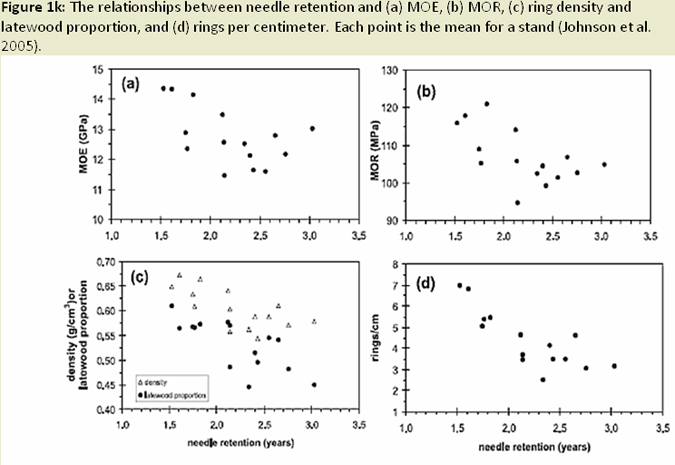
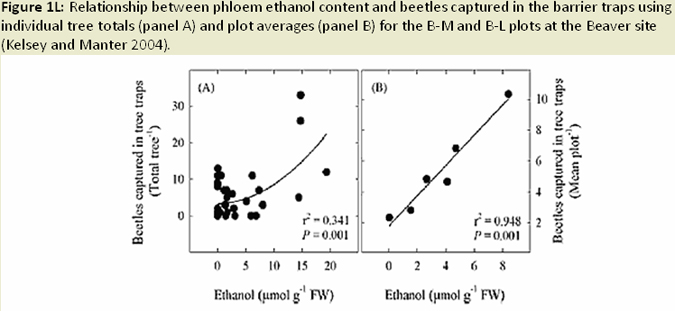
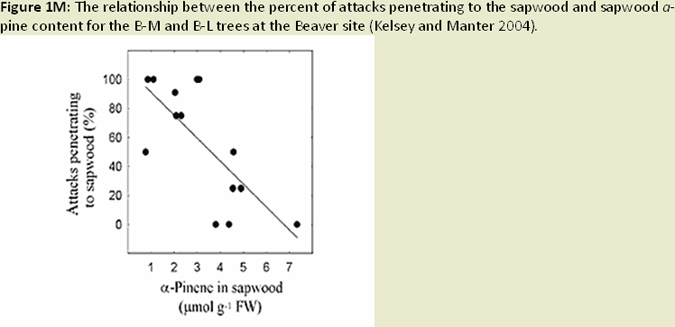
The distribution of foliage mass can be altered in infected trees, with the youngest age class foliage shifting towards the upper crown; while older age foliage shifted toward the lower crown (Weiskittel et al. 2006; Fig. 1I). Reduced foliage retention in upper crown corresponds to earlier findings regarding excess absorbed light and needle abscission (Manter 2002). Both leaf area and wood surface area decrease with increased SNC severity (Weiskittel and Maguire 2007). Reductions in leaf area, sapwood area, and permeability (Manter and Kavanagh 2003) relate to decreases in hydraulic conductance (Manter and Kavanagh 2003; Fig. 1J). In heavily infected stands, wood density and latewood proportion are much higher (Johnson et al. 2005) and annual growth (ring width) decreases, a result of reduced photosynethic capacity when needles are premature shed (Fig. 1K). Finally, ethanol concentrations (possible beetle attractant), wound-induced resin flow, and beetle attraction were all reduced as SNC severity increased (Fig. 1L). The success of attacks (penetration depth and gallery length) increased, most likely due to weakened defense mechanisms (oleoresin defense) of the tree (Kelsey and Manter 2004; Fig. 1M).